
Euvarroa sinhai is a mite that can infest the Western honey bee (Apis mellifera), yet our knowledge of it remains limited. Its original host is the dwarf honey bee Apis florea, which inhabits the subtropical and tropical zones of Asia. Beekeepers’ colonies are only occasionally infested by the mite, typically when A. florea populations are present in the same region. In the future, this mite may adapt to A. mellifera on a broader scale, posing a concern for future generations of beekeepers. Besides, it is not unusual for honey bee parasites to originate from other species, initially with localized impact, only to later spread globally. This review will examine the origin, biology, and ecology of E. sinhai, and evaluate its potential threat to apiculture.
Managed honey bee colonies, particularly those in intensive apiculture, are constantly exposed to parasites from other bee species. This is inevitable, as it is hard to find a place on earth where A. mellifera has not been introduced. In most cases of cross-species infestation, the parasite fails to complete its reproductive cycle and eventually disappears. Rarely, however, a parasite may undergo a host shift, establishing itself in the new host species. This phenomenon has been responsible for several significant epizooties, the most notable involving Varroa spp. and Tropilaelaps.

Image 1: Spread of Apis florea and Euvarroa sinhai. © Sémhur / Wikimedia Commons / CC-BY-SA-4.0
Infestation by E. sinhai occurs where imported A. mellifera colonies coexist with A. florea populations, which have expanded beyond their natural range and become invasive [1]. The mere presence of E. sinhai in A. mellifera colonies does mean there is a host shift, but the small risk remains; should this occur, it could trigger yet another epizooty in the Western honey bee.
Morphology and Biology
E. sinhai was first described in the international literature in 1974 by Mercedes Delfinado and Edward Baker, a couple of entomologists/acarologists. Females measure approximately 1.0 × 1.0 mm—about half the size of Varroa—with a densely hairy dorsal side and distinctive “whip-like” setae [2], [4]. Both Varroa and Euvarroa belong to the family Varroidae. The specific epithet sinhai honors Indian entomologist Ranendra Sinha [3].
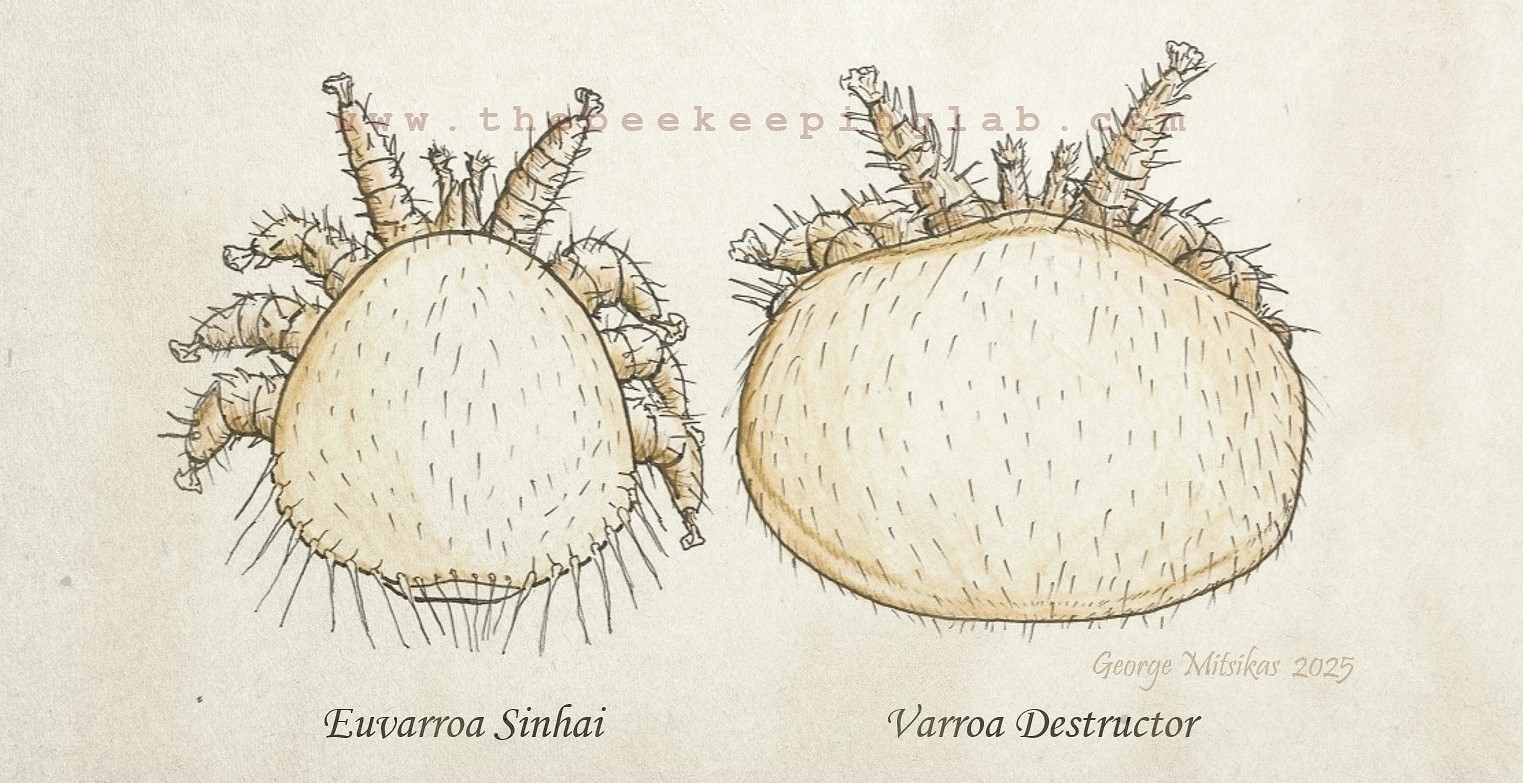
Image 2: Illustration of 2 mature female mites, Euvarroa and Varroa. The photographic material from Euvarroa is rare. For copyright reasons, photos are not published.
This mite is an obligate parasite of honey bees, requiring brood and adult bees for survival and reproduction. Its reproductive cycle in A. florea drone brood is described as “similar to Varroa jacobsoni” [4]. Natural parasitism occurs predominantly in drone brood [2], [4], [5], and is very rarely observed in worker brood [4], [5]. In the absence of drone brood, the mite remains phoretic on adult workers for extended periods. In adult A. florea, E. sinhai attaches to different abdominal segments than Varroa, producing a small but distinct protrusion [6]. As with Varroa, the adult female mite feeds on haemolymph and fat body tissue while on the host. It defecates after feeding, leaving conspicuous white spots at the feeding site [6], unlike Varroa, which defecates inside sealed brood cells. Laboratory studies have shown E. sinhai survival rates of 53% on adult A. mellifera, compared to 70% on A. florea and only 8% on Apis cerana [6].
Origin and Transmission
The red dwarf honey bees are the natural host of E. sinhai. A. florea has extended beyond its native range and is now considered invasive, with its presence documented as far afield as Malta in 2024 [9]. It is a non-managed species, nesting in the open on a single comb; however, its honeycomb is harvested and sold in several Asian markets. Transmission of E. sinhai to A. mellifera likely occurs via robbing behaviour and shared floral resources [7]. Apis florea has a high robbing tendency.

Image 3: Apis florea. This species usually nest around a shady branch in which they built a signle comb, but in a way they can perceive the position of the sun. Here they nest under the shade of a rock. [8]
Infestations in A. mellifera colonies are typically at extremely low levels, often involving only a few individual mites. To provide a point of reference, in an average conventional hive in an area where the Apis Florea colonies are native, natural fall traps caught one mite approximately every 23 days [7]. The mite has not been studied in areas where Apis Florea has invaded, and its status is unknown.
Phylogeny and Taxonomic Considerations
Euvarroa and Varroa share a common ancestor, likely parasitizing Asian honey bees during the Late Miocene (11.6–5.3 million years ago). This inference is deduced by paleomelittological evidence [10] and the phylogenetic relationships among extant species. Parallel with the geographic separation and speciation of honey bees, the Varroidae mites may have similarly diverged. Intriguingly, the giant honey bees (Apis dorsata, A. laboriosa, etc.) do not have their own varroidae mites—a gap possibly explained by displacement through Tropilaelaps spp. infestations [11].
The extant species of Varroidae and their authentic hosts are listed in the table below. The origin of Varroa Underwoodi is unknown, due to its wide geographical distribution.
| Genus | Species | Original Host |
|---|---|---|
| Varroa | V. Destructor | Apis Cerana |
| V. Jacobsoni | Apis Cerana | |
| V. Underwoodi | A. Cerana, A. Nigrocincta, A. Nuluensis | |
| V. Rindereri | Apis Koschevnikovi | |
| Euvarroa | E. Sinhai | Apis Florea |
| E. Underwoodi | Apis Andreniformis |
A second hypothesis could place the infestation of honey-producing bees of Southeast Asia by the ancestor of varroa mites chronologically later, after the diversion of the honey bee species. Such a scenario presupposes many host shifts. In any case, this must have occurred hundreds of thousands of years ago, perhaps even longer.
Potential Future Threats
As mentioned, E. sinhai is occasionally detected in A. mellifera colonies in Asia [2], but several barriers appear to limit its spread. The central question is whether it can successfully undergo a host shift to A. mellifera. Acarologists Kavinseksan and Wongsiri report no evidence of E. sinhai reproduction in A. mellifera brood under natural conditions [7], noting the absence of mites upon uncapping brood cells. However, laboratory studies confirm that E. sinhai can reproduce in A. mellifera brood [12], keeping the risk theoretical but plausible.
Interest in E. sinhai peaked in the 1980s–1990s, amid broader fears that all parasites of wild honey bee species could eventually adapt to A. mellifera. While these fears proved justified—given the number of documented parasite host shifts—E. sinhai faded from focus, likely because it never caused problems in apiculture.
For Varroa destructor, host shifts have been documented twice [13], [14], [15], with an additional third shift occurring possibly in the Vladivostok region of Russia (Primorsky Krai) around 1860. That early event did not produce an epizooty but resulted in A. mellifera populations with notable Varroa resistance, likely due to geographic isolation. Another Varroa species, V. jacobsoni, shifted hosts in Papua New Guinea [16]. Additionally, two Tropilaelaps species are known to parasitize A. mellifera brood. The main issue with all these host changes is that no one detected them at their onset, and therefore no one could observe the phenomenon in its development. Initial field identifications, in fact, were misguided both topologically and taxonomically due to the limited research means of the time. In other words, they did not identify neither the sources of each epizooty nor the correct parasite species. Some shifts may have resulted from small founder populations adapting to the new host; others from sustained spillover events. The role of human activity in facilitating such shifts remains obscure.
To summarize the risks, three major factors include the confirmed ability of E. sinhai to reproduce in A. mellifera brood (in laboratory settings), the invasive expansion of A. florea, and the latter’s strong robbing behavior. Mitigating factors include E. sinhai’s high specialization for A. florea, its minimal affinity for worker brood, and the absence of evolutionary pressure toward a host shift. Another potentially positive factor—despite the inherent risks—is interspecific competition with Varroa and Tropilaelaps mites [17], which may limit E. sinhai proliferation. Nevertheless, double or triple infestations cannot be ruled out. It is notable that A. mellifera and A. florea have coexisted in Southeast Asia for over 75 years, following the large-scale introduction of A. mellifera into the region, without E. sinhai becoming a problem for beekeepers. This suggests that, for now, the parasite remains stable within its original host niche. While this is reassuring, it cannot guarantee a benign future.
George Mitsikas
August 20, 2025
**Reposting of the article (reblogging, etc.), reproduction (in full/part), and use – reproduction of the photographs without the author’s permission is not allowed. Sharing the link on social media is permitted.**
REFERENCES
- El-Niweiri MAA, Moritz RFA, Lattorff HMG. The Invasion of the Dwarf Honeybee, Apis florea, along the River Nile in Sudan. Insects. 2019 Nov 15;10(11):405. doi: 10.3390/insects10110405. PMID: 31731633; PMCID: PMC6920986.
- Kitprasert, C. (1994). Parasitism by the brood mite, Euvarroa sinhai Delfinado and Baker (Acari: Varroidae) on the dwarf honey bee, Apis florea F.(Hymenoptera: Apidae) in Thailand.
- Presumably given by Delfinado and Baker as they did in Tetranychus Sinhai species.
- Akratanakul, P. (1975). Biology and systematics of bee mites of the family Varroidae (Acari: Mesostigmata).
- Kapil, R. P., & Aggarwal, K. (1989). OBSERVATIONS ON REPRODUCTION AND SEASONAL POPULATION TRENDS OF EUVARROA SINHAI (MESOSTIGMATA: VARROIDAE) IN INDIA. Progress in acarology, 2, 277.
- Koeniger, N., Koeniger, G., De Guzman, L. I., & Lekprayoon, C. (1993). Survival of Euvarroa sinhai Delfinado and Baker (Acari, Varroidae) on workers of Apis cerana Fabr, Apis florea Fabr and Apis mellifera L in cages. Apidologie, 24(4), 403-410.
- Kavinseksan, B., & Wongsiri, S. (2016). Grooming behavior of Apis dorsata Fabricius, Thai commercial, and Primorsky honey bees (Apis mellifera Linnaeus) to the bee mite Euvarroa sinhai Delfinado & Baker. Journal of Asia-Pacific Entomology, 19(2), 359-363.
- By Persium – Own work, CC BY-SA 4.0, https://commons.wikimedia.org/w/index.php?curid=155888302
- Uzunov, A., Galea, T., Chen, C., Cilia, G., Costa, C., & Mifsud, D. (2024). Apis florea in Europe: first report of the dwarf honey bee in Malta. Journal of Apicultural Research, 63(5), 1122–1125. https://doi.org/10.1080/00218839.2024.2386888
- Greater past disparity and diversity hints at ancient migrations of European honey bee lineages into Africa and Asia. Journal of Biogeography, 40(10), 1832-1838.
- Euvarroa mites are found under Apis Dorsata nests, but they are a result of contamination.
- Mossadegh, M. S. (1990). Development of Euvarroa sinhai (Acarina: Mesostigmata), a parasitic mite of Apis florea, on A. mellifera worker brood. Experimental & applied acarology, 9(1), 73-78.
- Anderson, D. L., & Trueman, J. W. H. (2000). Varroa jacobsoni (Acari: Varroidae) is more than one species. Experimental & applied acarology, 24(3), 165-189.
- Navajas, M., Anderson, D. L., De Guzman, L. I., Huang, Z. Y., Clement, J., Zhou, T., & Le Conte, Y. (2010). New Asian types of Varroa destructor: a potential new threat for world apiculture. Apidologie, 41(2), 181-193.
- Two host shifts are known, one fairy old in Japan in the end of the 19th Century and another one in Korea or somewhere in the North. Kaise, S., & Hoshiba, H. (2023). Revisiting the History of Parasite Conversion of Varroa destructor to Apis mellifera–the Parasitic Shift Occurred as Early as 1898?. Bee World, 100(3-4), 71-74.
- Roberts, J. M. K., Anderson, D. L., & Tay, W. T. (2015). Multiple host shifts by the emerging honeybee parasite, Varroa jacobsoni. Molecular Ecology, 24(10), 2379-2391.
- Sammataro, D., Gerson, U., & Needham, G. (2000). Parasitic mites of honey bees: life history, implications, and impact. Annual review of entomology, 45(1), 519-548.

Leave a comment