
Honey bees stand out from other bee species due to three characteristics. The first is polyandry, that is, the mating of the queen with several drones. The second is genetic diversity, which ensures that the colony expresses no rigid behaviors. The third is something so obvious that it is overlooked: Iteroparity. The colony is a multitude of offsprings, and its queen a “machine gun” of repeated births. In this article, we develop iteroparity and its importance, as well as the problems associated with it.
Animal organisms have different reproductive strategies and are divided into semelparous, iteroparous and intermediates 1. Semelparous species have one reproductive episode, usually terminal (e.g. Cicadas), while iteroparous species have repeated births for life. As for the bees, solitary bees tend to semelparity, and social ones to iteroparity. Semi-social bees (bumblebees, etc.) have a mix of two motifs due to the great competition between the females of the colony. In this text we focus on Apis Mellifera which is clearly iteroparous, both individually (lays workers and drones), and as a superorganism. In each swarming event the colony is divided, the old queen departs, and offspring swarms are formed. A queen bee can live more than four years when it is found in a colony that is in its natural habitat. During this time it will swarm several times and produce many new colonies 2.
Superorganisms and iteroparity
Iteroparity has emerged through the evolutionary course of Apis Mellifera, whose ancestral state was the semi-social. Due to a behavior within the nest called “policing”, their species gradually evolved into a fully social one. Thousands of years of evolution favored the differentiation between the female individuals of each colony. There was a point of inflection at which the queen ceased to give birth to full worker-bodied females. Beyond that stage, there was no return to the previous physiology. This is how castes and superorganisms were formed.

Generally in nature, births and parenting require a large expenditure of energy, but not in superorganisms. In this case the main energy barriers are removed because castes exist. Thus, the somatic preservation and the production of offsprings no longer limit each other, as it happens in multicellular organisms. In superorganisms there is so much specialization that iteroparity is extreme as far as sterile workers are concerned.
Would anyone wonder why the beehive appears so “wasteful” when there is the so-called “economy of nature”? Why honey bees raise so many swarming cells when he could bring out two or three? The colony does it, after all, when queens are replaced. The answer is that the above queen cells are not a waste of energy. The energy is saved by the limited care for offspring swarms (rather than individual cells). The energy favor is always given to the maternal swarm, and the “parental investment” is small for each new queen. Daughter swarms are generally smaller than their mother’s.
The parameters of Iteroparity
The iteroparity of a hive that does not receive major interventions can be estimated according to three parameters:
- The frequency/tendency of swarming.
- The average number of swarming cells (queen descendants)
- The tendency to lay/ rearing workers.
All three factors are genetically predetermined. The extent to which they will be expressed, however, depends on the environmental conditions, the size of the swarm, beekeeping manipulations etc. The degree of iteroparity is something of a constant and is related to the reproductive instinct of each locally adapted bee (ecotype). It has nothing to do with whether a queen lays a lot, lays spotty brood etc. Each ecotype has its own parameters of iteroparity.
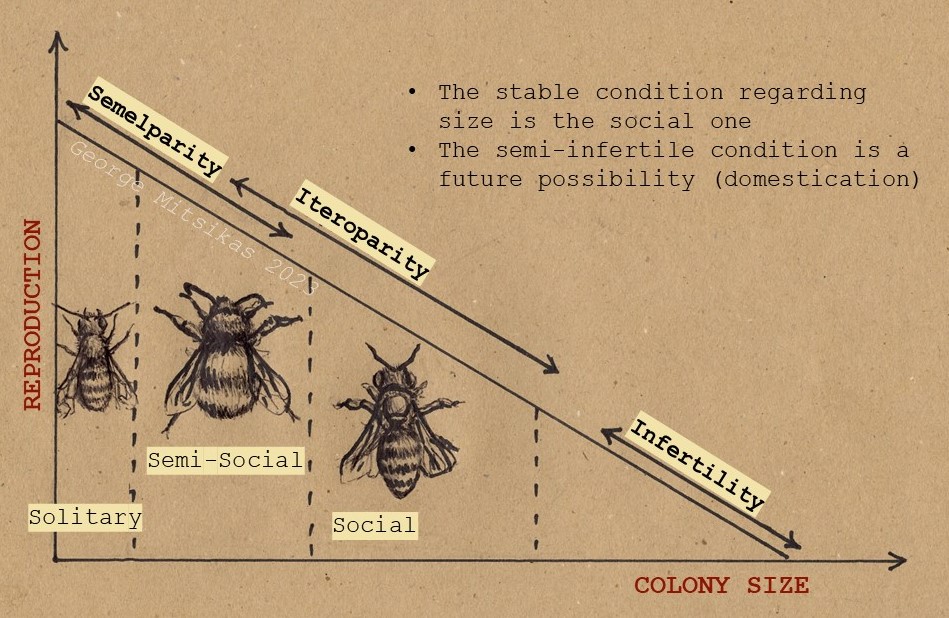
When worker rearing increases a lot, then the hive works against its own iteroparity principle. The reason is that the creation of very large colonies is favored during the year without a reproductive episode. Classical genetic improvement struggles to increase this very parameter, because this ensures larger honey crops. The problem arises when this venture is combined with attempts to eliminate the instinct of swarming and enhance gentleness, which may entail ignoring the predators. The combination of these man-made efforts pushes the evolution of common bee towards domestication.
In the recent decades queens tend to fail close to two years of life. Bees replace them. This phenomenon, which has taken on a global dimension should be of concern, because it cancels out the iteroparity of the European bee. Iteroparity apparently assumes that the average queen can live more than 2 winters. The fact that apiaries are constantly acquiring many queens from replacement may lead to deviations from normal (see below).
The Last Survivor
The evolutionary value of iteroparity in social bees can be betterly understood through the classical beekeeping debate about “which queens are superior?“. Queens from swarming cells are often compared with queens from classical selective breeding. One portion claims that the swarming are superior and another that the grafted/selected are the superior. A third group considers artificial insemination queens the best. The paradox is that all three factions consider that the superiority of their version stems from the principle of selection! The first claim natural selection, the second claim man-made selection through experience and the third man-made selection through technology. Now, let’s consider the reproductive steps of the first version. Natural “selection”. According to this, the beehive during swarming enters a sequence of decisions and trials:
The beehive chooses when to swarm waiting for the right moment. Then the beehive chooses which of the queen cells to keep and seals them. After the maternal swarm leaves the hive, a struggle takes place as to which of the hatched sister queens will prevail, (before departure, in the decoctions with many queens, etc.) During mating flights we have another step of natural selection: Which drones will be strong enough to catch up with the queen? After mating, the queens should be able to avoid birds and other enemies and turn around. Then, another competition begins. Which of the survived swarms will be competent enough to settle? And so on…
That is, out of the hundreds of candidate “genetics” of each colony, only one will be left in the end. In the end there can be only one…

If each of the above stages was a true selection supernatural bees would have emerged within a few years! This reasoning is misleading and it is called “survivorship bias”. The cognitive fallacy in which supernatural abilities are attributed to individuals who survive an elimination process. Many of the above steps require skills that take out of the game any defective individuals, but they also require a lot of luck.
After the all defective individuals are rejected, another principle prevails: Equation. The superorganism does not make genetic distinctions between its own “children”. Equality, fraternity and diversity. On the contrary, the distinctions of descendants have a place in the previous evolutionary stage, the semi-social. The main distinction of which larva will become queen is the nutritional status of the larva 3. In the long term, iteroparity exists so that the bees do not have to make any other choice than the nutritional.
The reason why queen rearing from swarming cells, as well as from grafting are so successful, lies in the fact that there is no discrimination regarding a specific parameter: Both methods do not favor the so-called “royal lines”. Royal lines are specific larvae that emit chemical signals for nurse bees to favor them as queens 3. This phenomenon, which occurs during queen replacement and during emergency queen rearing, often works against the principles of altruism and polyandry. It is a small setback to the ancestral, semi social state.

The colony overcomes the problem of usurper “royal” larvas by making the so-called “cups”. They are forerunners of reserve queen cells waiting for the queen to lay an egg in them in a random way and thus not to favor some royal line. But this is not always the case in replacement. The large hives which are orphaned at the heart of spring may be the exception. They probably produce random queen cells. To wrap up, the continuous production of colonies via emergency cells and the frequent failing of queens favors the cryptic royal lines and “awakens” semi-parasitic, or primordial behavior 4 ,5 . Unfortunately, the books and the internet made popular the methods of emergency queen rearing, which are often followed under the pressure to increase the number of beehives.
As for the average superiority of queen from swarming cells over the average grafted queens, this can be attributed to epigenetic factors, as we described in a previous article. We should take into account that colonies in a swarming state are in no hurry to rear queen cells, while they breed them under optimal conditions. Unlike most grafting techniques in which the bees work in a state of orphanage, in large crowds and not always at the hart of spring. In the face of these disadvantages, classic breeding counteroffers the element of human selection and so the great debate continues on and on…

Iteroparity and Randomness
To see how the system of randomness works during swarming, here is a comment by the old beekeeper Dave Cushman 6 :
…”The idea that swarming is no just the reproduction of honey bee colonies on a colony by colony basis, but that there is also an element of exploration of forage and suitability of conditions in areas that are adjacent to the swarming colonies original location […]. I noticed that some colonies would swarm to such an extent that the parent hive did not survive. I am of the opinion that bees do nothing inadvertently and I puzzled over what their (nature’s) strategy might be. The conclusion that I came to stemmed from the numbers of swarms that issued and the directions of their escape […]. The different directions suggested to me that the bees swarming at the time were aware of the existence of the freshly established swarms that had left hours or days before. And that they did not want to compete with them. It then became plain to me that the bees were not ‘propagating the race’, but were exploring the geography. Any swarms finding a good place would thrive whilst those in poorer areas would fail.”…
With this natural system, the identification of more suitable places is favored in the long run, and at the same time maladaptive bee phenotypes disappear. Essentially natural selection in Apis Mellifera is done through exclusion. It is a negative selection process (nature tends to reject the maladaptive). On the contrary, classical breeding/ grafting is positive selection. In the case where nature makes a positive choice on its own, the great tendency to swarming is favored, which is not in our interest. The evolutionary advantage of iteroparity is that while survival roulette rarely wins, Apis Mellifera (her cousin Apis Cerana etc.) can play over and over again. We do not buy the widespread view that “nature never leaves things to chance“. It is anti-Darwinian. Same goes for the cliché of the “survival of the fittest”.
Ecotypes 7
Globally, the parameters of iteroparity have been strongly influenced by humans. For instance, an ecotype that swarms easily may be attributed to the fact that bees were kept in coffins for many centuries (not to a physical adaptation). The large size of the colonies is another feature that was reinforced by humans. The different physiological parameters of all ecotypes can be evident in overwintering. Some ecotypes favor the individual fat body and some others favor the number of individuals. Eg. Native northern honeybees may have a smaller critical mass in their winter clusters than bees under the Mediterranean climate. Conversely, the fat body of the Northen honeybee has a hypertrophic tendency. The diapause of the queen and the decrease in the activity of the colony in fall are decisive for its development.

The successful beekeeper must therefore be able to sketch the profile of the local bee. To maintain bees that align with the ecotype rule. Beekeepers must therefore leave behind a desire for very large colonies when their ecotype does not “allow it”.
To sum up, we have seen how the stability of the European honey bee is currently under threat. Semi-domestication, short-lived queens, poor methods of multiplication and ignorance of ecotypes. Everything somehow relates to the oversight of the basic “trinity”: Polyandry-Diversity-Iteroparity. The old beekeepers claim that the old bees were way better than current ones in performance. We believe this happens not only due to varroasis and environmental factors, but also due to the contribution of the above degenerative factors.
George Mitsikas (chemist, amateur beekeeper)
March 2023
*It is not allowed to republish the article (reblogging, etc.), or reproduce it (total / partial) without the permission of the authors.*
NOTES
- Here the word “strategy” is a metaphor to describe the adaptive patterns that emerged through the evolutionary process of Apis Mellifera. I claim that biological processes are essentially contingent and definitely non-teleological (non-purposeful).
- Tom Seeley estimates that a colony in the wild lives up to five years. After that time its nest cavity is usually occupied by another swarm. The lifespan of the older queen (foundress) however, cannot be easily estimated.
- This happens under emergency queen rearing, but in principle is true. See Sagili, R. R., Metz, B. N., Lucas, H. M., Chakrabarti, P., & Breece, C. R. (2018). Honey bees consider larval nutritional status rather than genetic relatedness when selecting larvae for emergency queen rearing. Scientific Reports, 8(1), 1-11.
- Withrow, J. M., & Tarpy, D. R. (2018). Cryptic “royal” subfamilies in honey bee (Apis mellifera) colonies. PLoS One, 13(7), e0199124.
- More about Apis Mellifera mutiny behaviours etc one may find in the lectures of entomologist Keith Delaplane https://www.youtube.com/watch?v=Wu-XlsJie3A
- http://www.dave-cushman.net/bee/sgecf.html
- The notion of the ecotype describes better the “locally adapted bee” as the old bee races have been intermixed.

Leave a comment